��ѯ����: 400-670-9909
��ϵ����
ϸ������֯�������ܹ��ɹ������������������������Ż���һ������ͼģ�����ڵ������������ü�����һ���ؼ���������������ѡ����������ͨ�������Ӷ���Ѫ�塣Ѫ���������ɷ���ɵĸ߶ȸ��ӵĻ�����Щ�ɷְ����������ӡ����ء���Ԫ���Լ�ճ������չ���ӵȣ����Ƿ����Ŷ�������ѧ���ԡ�
Ѫ���а��ĺ����ߣ���������ϸ��״̬��ά�֣����û��ư�����ֹpHֵ���������Ƶ���ø���Ժͱ���ϸ�����ܼ��������˺���[1,2]������Ѫ����Ժܺõ�ά��ϸ������֯��������������Ϊ������������������Ե�ȱ�㣬��ɱ��ϸߡ���Դ���١�ʵ������Դ���Ӱ�����β����ʹ���DZ�ڵ���ȾԴ�Ͳ������صȡ����⣬���ǶԶ��ﱣ����ʶ�����Ҳ������Ѫ���ʹ��[2, 3]��
Ѫ����ϸ������Ӧ���е���Щȱ��ʹ�ý�Щ���ڿ�ѧ�о���ҵ�����������Ƕ���Ѫ����������������������[3]��
�����ܹ��˷�Ѫ���ȱ�㣬��Ѫ������������������ͻ�����ŵ㣬��������ض���ϸ����������ר�õ����������Լ��ɾ�ȷ�ؿ���ϸ������ֳ�ͷֻ�����[2]��
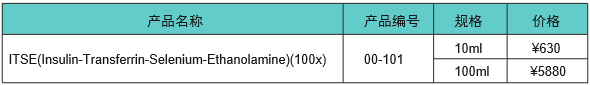
��������Ѫ����������ȻӪ���ḻ���������ṩϸ���������Ӫ���ɷ֣���ͨ������Ҫ��һ�������ض�����֣����������ӡ�֬���ᡢά���غ���Ԫ�صȡ������ʵ�����������ڴ����ϸ������Ѫ�����������ڻ���������(��ΪDMEM/F12)�б����������ITS(�ȵ���-ת������-��)[4]��������ijЩϸ�������������л�����Ҫ�����Ҵ���[5]��
Ϊʲôѡ��ITSE?
ѡ��ITSE���������ͨ�����õ�ITS���������ڣ��û��Լ�������ʶ��������������ϸ����Ҫ�Ҵ������ܴﵽ��ѵ�����״̬��ʹ��ITSE������ɱ�֤�������͵�ϸ�����ܵõ������Ӫ����Ӧ��
�ȵ��� (Insulin)
�ɴٽ�ϸ������������ϸ���������ǡ��������֬�����ȡ�����á��ȵ���Ҳ���еֿ�ϸ�����������á��ڲ��鶯��ϸ������Ѫ������ʱ�������ȵ��ص�����Ũ��Զ����������Ũ�ȣ������ȵ��س������ȵ�������(Insulin Receptor, IR)��ϣ����Ӵ�л���ںͿ����������⣬����ͨ�������ȵ�������������(IGF-I)�����壬���Ӵ���˿���ѺͿ��������ã�������������ЧӦ[6]��
ת������ (Transferrin)
��Ѫ���и�����һ���ǵ��ף��������������ӿ����Խ�ϡ����������ӵ�ת�˺ʹ�л�з�����������Ҫ�����ã�ͬʱҲ��һ����Ҫ�İ��⿹������ ����ˣ�ת����������Ѫ���������ز����ٵ������ת����������Ǵ�Ѫ���з�������ģ�Ȼ������������Ͱ�ȫ�Բ�����⣬�����ת�����ױ�Խ��Խ�㷺��Ӧ�á��Ӹ��ֱ���ϵͳ������ֲ���л�õ�����ת�������������ƵĻ�[7]��
�� (Selenium)
��һ����Ԫ�أ����ǹ����Ĺ�������ø�� ����������ԭø��������������ø�Ĺؼ���֡� ���ѱ�ȷ������Ѫ���������ı���������[8]��
��һ���л�����������Dz���Ҳ�Dz���������Ϊ��֬�ϳɵ�ǰ�塣�Ҵ������ӽ���ϸ������ֳ�з�������Ҫ�����ã�����������ӵ��ӽ���ϸ������Ѫ�������С��Ҵ���Ҳ������������ϸ������Ѫ������[5, 9]��
�ο����ף�
1. Brunner, D., Frank, J., Appl, H., et al. (2010). Serum-free cell culture: the serum-free media interactive online database. Altex, 27(1), 53.
2. Freshney, R. I. (2005). Serum�\Free Media. Chapter 10 in: Culture of animal cells, John Wiley & Sons. pp, 129-143.
3. Butler, M. (2013). Serum-free media: standardizing cell culture system. Pharmaceutical Bioprocessing, 1(4), 315-318.
4. Gstraunthaler, G. (2003). Alternatives to the use of fetal bovine serum: serum-free cell culture. Altex, 20(4), 275-281. 5. Murakami, H., Masui, H., Sato, G. H., et al. (1982). Growth of hybridoma cells in serum-free medium: ethanolamine is an essential component. Proceedings of the National Academy of Sciences, 79(4), 1158-1162.
6. Kim, J. J., & Accili, D. (2002). Signalling through IGF-I and insulin receptors: where is the specificity?. Growth Hormone & IGF Research, 12(2), 84-90.
7. Brandsma, M. E., Jevnikar, A. M., & Ma, S. (2011). Recombinant human transferrin: beyond iron binding and transport. Biotechnology advances, 29(2), 230-238.
8. Saito, Y., Yoshida, Y., Akazawa, T., Takahashi, K., & Niki, E. (2003). Cell death caused by selenium deficiency and protective effect of antioxidants. Journal of Biological Chemistry, 278(41), 39428-39434.
9. Tsao, M. C., Walthall, B. J., & Ham, R. G. (1982). Clonal growth of normal human epidermal keratinocytes in a defined medium. Journal of cellular physiology, 110(2), 219-229.





























 ���������� 31011202004777�� ����֧�֣���������
���������� 31011202004777�� ����֧�֣���������