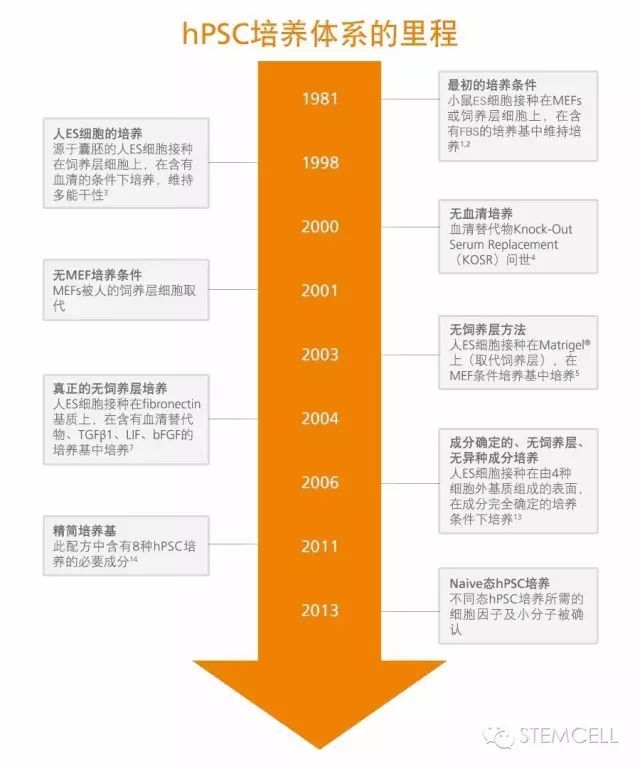
���ܸ�ϸ��������ϵ����ʷ����MEFs��Naïve State
��������hPSCs�����Ľ���
������̥��ϸ����hESCs�������������������������˶��ܸ�ϸ����hPSCs���������������ϵı����������1,2�����������������ϸ����Ҫ������С����̥����άϸ����mouse embryonic fibroblasts, MEFs������������ϸ�������Ϻ�̥ţѪ�壨FBS����������3�����ǣ����ܸ�ϸ�����ٴ�Ӧ��ǰ��ʹ���Ǻܿ���ʶ�����������в��ɴ�����Щ����DZ������ԭ�ԵĶ���Դ�ɷ֣��������hPSC�����ķ������������������������в�ȷ���ĺͷ���Դ�ijɷ���4-6��������ͼ��ʾ��hPSC���������ݱ�����������㣬�������������������ӣ�bFGF ��/�� TGF-�£���Ѫ��������ʹ�ð������Matrigel������������7,8���������������Ҳ��̷����˳ɷ�ȷ���ģ��������㣬�����ֳɷ֣�����������ɷ֣���hPSC�������䷽��9-12����

���������������� vs ��������
2006�꣬Dr. James Thomsonʵ���ҵ�Dr. Tenneille Ludwig�Լ�WiCell Research Institute��Madison, WI����ѧ���Ƿ������ڳɷ���ȫȷ�����������㲢�����ֳɷֵ������������µ�ESϸ��ϵ��12, 13�������ǽ����bFGF��ά��PSC�����Ҹ�����������TGF-�£�����PSC�ķֻ�����һϵ����������Ҫ���ӣ�����PSC�Ķ����ԣ�ʹhPSC�������������������ʵķ�Խ�����䷽�ĸ����� - mTeSR1�Ѿ���Ϊ�������������Ľ������ֹ��2016�꣬mTeSR1�����ó���1100�Ρ�
2011�꣬Dr. James Thomsonʵ���ҵ� Dr. Guokai Chen������һ�ֵ͵��ף��������hPSC�������䷽��������ΪE8��14����TeSR-E8 ��E8�䷽������hPSC����8�ֱ�Ҫ�ijɷ֣�insulin, selenium, transferrin, L-ascorbic acid, bFGF��TGF�£�NaHCO3��DMEM/F12��14����
ϸ�����ڻ��ʣ�Matrigel vs Vitronectin
Matrigel��һ��Դ��Engelbreth-Holm-Swarm��EHS��С������ϸ���Ļ���Ĥ���ʣ���mTeSR1 ���ʹ����hPSCs��������12������Dr. Tenneille Ludwig������Ҳ��һ�ֲ�������ԭ�Ե���DZ����Դ����������о����������ҵ���Matrigel���ӳɷ�ȷ���Ļ��ʡ����бȽ���ǰ������ͨ����ѧ���ӵ��������ϵ��˹�������15,16��������ij��integrins��ϸ��ճ�����ӽ�ϵ����鵰����17,18������Primorigen Biosciences��˾���ƿ�����Vitronectin XF����һ�ֳɷ�ȷ���������ֳɷֵĻ��ʣ��������Matrigel֧��hPSC��ά��������Vitronectin XF��TeSR������������������ݣ���mTeSR1һ��ʹ�ôﵽ�ɷ�ȷ��������ϵͳ������TeSR2��TeSR-E8������һ��ʹ��ʱ���ﵽ��ȫ�����ֳɷֵ�����ϵͳ��
ϸ���������ſ� vs ��ϸ��
��ͳ��hPSC������������ֱ��Ϊ50 �C 200��m���ſ���д��������ִ����������������ڿ����ڶ�δ�����>20�������Ա���ϸ���ĺ��͡�hPSCҲ�ɽ��е�ϸ���������������ױ��������ַ������ϸ����ɲ���Ҫ��ѡ��ѹ���Ӷ����º��ı䣬��˲����Ƽ���19, 20����ʹ�õ�ϸ������ʱ��Ӧ���ڽ��м���Ա�֤�����ȶ���
ϸ��������ø vs ��ø
����������������������hPSC��ͨ���β���/��ø�����ɢø�������������ſ����д��������Ǵ˹���ʮ�ֺ�ʱ�����ſ�Ĵ�С���ѿ��ơ���Ȼø�����Ĺ�Ч��������ø������ϸ���⵰�ף�����ϸ�������ˣ�����Ҫ����ϸ��֮ǰʹ��ʧ�
��ø�����������º�ϸ������Һ��Gentle Cell Dissociation Reagent, GCDR���ɴ�Ӧ�˶�����GCDR�����ƻ�ϸ�����浰�ף��������ʧ�������ϸ��ʮ���ºͣ�����ʹ�������������ߡ�
2015�꣬һ��������ø��������ReLeSR�����ˡ������ֹ��β���ReLeSR�ɽ�δ�ֻ���hPSC���ſ����ʽ���������Ϸ��룬���ֻ���ϸ���Ա������������ϡ����ִ�����ʽ����ʡ��ʱ�䣬���ҿ��������ܷ�������ϸ��������
���ģhPSC������2D vs 3D
hPSC��ҩ��ɸѡ�����Լ���ϸ�����������е�Ӧ����Ҫ��������ʼϸ����ijЩ�ٴ�Ӧ�������ϸ�����Ѿ����ij�����2D�����ܹ��ṩ��ϸ�����������3D��������Ӧ�˶�������3D���������У�hPSCs���Ը����������ϻ������ſ����ʽ��������������������������ҡƿ����ͽ������ﷴӦ����21-25����hPSC����������Ҫ��������������Ҫ��ͬ�� ���磬�����������У�hPSC���Բ��û��ʣ��Ծۼ���ķ�������������������2D���������ﵽ�ġ����⣬�ڴ��ģ���ﷴӦ���н��ϲ��ԣ�����ע�Է������ϣ���Ϊ��Ҫ�� ��ˣ������о��Զ����ڴ��ģhPSC�����ķ��������Ż�������߲������ȶ��Ժ�Ч�ʡ���2016��ISSCR�����ϣ���һ������hPSCs����������������ӳ�����ǵ����� - mTeSR3D��ͨ���Ż��䷽�ͷ������ϵ�����������mTeSR3D�ṩһ�����Լ۱ȵķ��������ڶ̶�2-3��֮�ڣ�������1x109ϸ����
��̬ͬhPSC������Naïve-like vs Primed
��ͳ�����ϵ�hPSCs����Ϊ���ڡ�primed state�� ����С��ֲ������߲��ϸ����EpiSCs��������26�������֮�£�С���ESϸ������Ϊ����naïve��ground state�������и��ԡ������ ���ʵ���ұȽ���С��ESϸ���ʹ�ͳhPSCs������������ʾ����ͬ�Ļ������ģʽ�����Ҷ�����������Ҫ��Ҳ��ͬ��27-29����������˵��ά��δ�ֻ�״̬��С��ESϸ����Ҫ������Ѫ���������ӣ�LIF����������������hPSC��С��EpiSC����FGF��Activin������������LIF����Ӧ27�����о�����primed state��hPSCs���Ա�ת�ص�ground state��28, 30-32��������һ��֧��hPSCs��naïve-like state��������Ҳ�Ѿ����� - RSeT��ʹ��mTeSR1��RSeT������������ʹhPSCs��primed state��naïve-like state֮�������л���
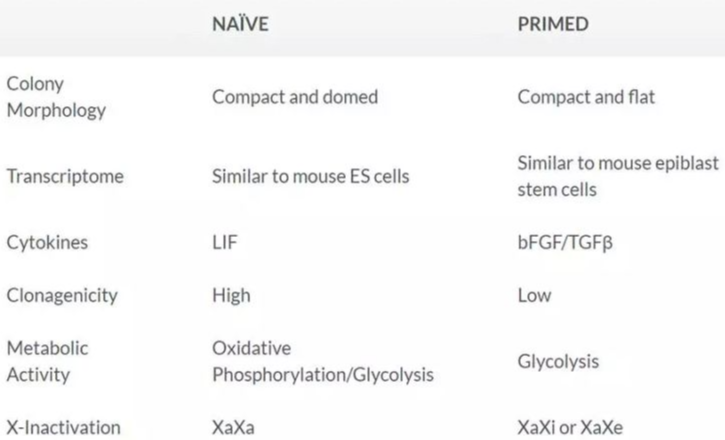
Table 1. naïve-like state��primed state���ܸ�ϸ���ıȽϡ�
������
hPSCs������ϵͳ�ڹ�ȥ�ļ�ʮ���������˴���ȵĽ�������ʹ��hPSCs�������Ӽ�������һ���ԣ�������������һ�ж�ʹhPSCs�����ƣ�ҩɸ�������е�Ӧ�ø���һ������������������ᵽ������ϵͳ����Ȥ�����⣬��ӭ����ע���ǵĹ����˺ţ���������ͨ���绰/�ʼ���ϵ��
References
1.Evans MJ & Kaufman MH. (1981) Establishment in culture of pluripotential cells from mouse embryos. Nature 292(5819): 154�C6.
2.Martin GR. (1981) Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci U S A 78(12): 7634�C8.
3.Thomson JA et al. (1998) Embryonic stem cell lines derived from human blastocysts. Science 282(5391): 1145�C7.
4.Amit M et al. (2000) Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Dev Biol 227(2): 271�C8.
5.Hovatta O et al. (2003) A culture system using human foreskin fibroblasts as feeder cells allows production of human embryonic stem cells. Hum Reprod 18(7): 1404�C9.
6.Xu C et al. (2001) Feeder-free growth of undifferentiated human embryonic stem cells. Nat Biotechnol 19(10): 971�C4.
7.Amit M et al. (2004) Feeder layer- and serum-free culture of human embryonic stem cells. Biol Reprod 70(3): 837�C845.
8.Levenstein ME et al. (2006) Basic fibroblast growth factor support of human embryonic stem cell self-renewal. Stem Cells 24(3): 568�C74.
9.Vallier L et al. (2005) Activindal and FGF pathways cooperate to maintain pluripotency of human embryonic stem cells. J Cell Sci 118(19): 4495�C509.
10.Lu J et al. (2006) Defined culture conditions of human embryonic stem cells. Proc Natl Acad Sci U S A 103(15): 5688�C93.
11.Yao S et al. (2006) Long-term self-renewal and directed differentiation of human embryonic stem cells in chemically defined conditions. Proc Natl Acad Sci U S A 103(18): 6907�C12.
12.Ludwig TE et al. (2006) Feeder-independent culture of human embryonic stem cells. Nat Methods 3(8): 637�C46.
13.Ludwig TE et al. (2006) Derivation of human embryonic stem cells in defined conditions. Nat Biotechnol 24(2): 185�C7.
14.Chen G et al. (2011) Chemically defined conditions for human iPSC derivation and culture. Nat Methods 8(5): 424�C9.
15.Melkoumian Z et al. (2010) Synthetic peptide-acrylate surfaces for long-term self-renewal and cardiomyocyte differentiation of human embryonic stem cells. Nat Biotechnol 28(6): 606�C10.
16.Villa-Diaz LG et al. (2010) Synthetic polymer coatings for long-term growth of human embryonic stem cells. Nat Biotechnol 28(6): 581�C3.
17.Rodin S et al. (2010) Long-term self-renewal of human pluripotent stem cells on human recombinant laminin-511. Nat Biotechnol 28(6): 611�C5.
18.Nagaoka M et al. (2006) E-cadherin-coated plates maintain pluripotent ES cells without colony formation. PLoS One 1: e15.
19.Draper JS et al. (2004) Recurrent gain of chromosomes 17q and 12 in cultured human embryonic stem cells. Nat Biotechnol 22(1): 53�C4.
20.Buzzard JJ et al. (2004) Karyotype of human ES cells during extended culture. Nat Biotechnol 22(4): 381�C2; author reply 382.
21.Olmer R et al. (2010) Long term expansion of undifferentiated human iPS and ES cells in suspension culture using a defined medium. Stem Cell Res 5(1): 51�C64.
22.Oh SKW et al. (2009) Long-term microcarrier suspension cultures of human embryonic stem cells. Stem Cell Res 2(3): 219�C230.
23.Singh H et al. (2010) Up-scaling single cell-inoculated suspension culture of human embryonic stem cells. Stem Cell Res 4(3): 165�C79.
24.Krawetz R et al. (2010) Large-Scale Expansion of Pluripotent Human Embryonic Stem Cells in Stirred-Suspension Bioreactors.
25.Zweigerdt R et al. (2011) Scalable expansion of human pluripotent stem cells in suspension culture. Nat Protoc 6(5): 689�C700.
26.Rossant J. (2008) Stem Cells and Early Lineage Development. Cell 132(4): 527�C531.
27.Nichols J et al. (1998) Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 95(3): 379�C391.
28.Takashima Y et al. (2014) Resetting Transcription Factor Control Circuitry toward Ground-State Pluripotency in Human. Cell 158(6): 1254�C1269.
29.Marks H et al. (2012) The transcriptional and epigenomic foundations of ground state pluripotency. Cell 149(3): 590�C604.
30.Gafni O et al. (2013) Derivation of novel human ground state naive pluripotent stem cells. Nature 504(7479): 282�C6.
31.Theunissen TW et al. (2014) Systematic Identification of Defined Conditions for Induction and Maintenance of Naive Human Pluripotency. Cell Stem Cell 15(4): 471�C487.
32.Chan YS et al. (2013) Induction of a human pluripotent state with distinct regulatory circuitry that resembles preimplantation epiblast. Cell Stem Cell 13(6): 663�C675.





























 ���������� 31011202004777�� ����֧�֣���������
���������� 31011202004777�� ����֧�֣���������